マイクロラーニング
隙間時間に少しずつビデオや記事で学べるマイクロラーニング。クイズに答えてポイントとコインを獲得すれば理解も深まります。

ヨガ療法における筋膜 パート1/2
ヨガは、揺れている認識の波の中で、その存在を明らかにすることをやめたマインドにおいて経験されるものである - パタンジャリのヨガ経典より, ca. 150 CE 1.2 (2002年スタイルスにて翻訳) 筋膜セラピーとしてのヨガ ヨガ(=絆、調和、対立とのバランス)は、先史時代のいつ頃か、格闘技関連分野に起源を持つ、心身的なセルフトレーニングの一種として生まれました。(Feuerstein, 1988). ヨガは今も昔も「空間的な医療」への主だった探究の形なのです - 人の形を変えることにより機能がどのように変化するのでしょうか? (Myers 1988) 上で引用されたヨガにおける最初の実践的な文章は、およそ2000年前に書かれたものです。 ヨガとは非常に複雑な自己認識システムです。そこにおける精神、感情、そしてスピリチュアルな領域の地図や解説には、ヒンドゥー教やアーユルベーダのヒーリングシステムにもあるような寓話的で潤沢なイメージが含まれています。(Lad 1984) 繊細さ、ヨガにおける「八正道」、チャクラ、そして瞑想状態といったものは有益なものではありますが、ここで取り扱う範疇をこえているので、ここではフィジカルトレーニングの「四肢」とも言えるハタヨガについてのみ言及していきます。 ゴール: ハタヨガがセラピーとして使用される場合、ゴールには以下の要素の向上が含まれます: ストレングス バランス スタミナ 柔軟性、そして リラクゼーション テクニック: 実践的なところでは、ハタヨガはクラス形式であったり、より細かく行う場合には1対1のヨガセラピーとして行われます。以下に記述する原則や方法はどちらの形式にもあてはまりますが、いくつかの1対1形式に特化された要素は後述します。ヨガセラピーの主なツールは以下のものを含みます: プラーナヤーマ(心を落ち着かせ、リラクゼーション反応を呼び起こし、自律的生理機能を向上させるためにデザインされた呼吸練習法) アーサナ(縮こまったり、固まった組織を連動/ストレッチ、弱い筋肉を強化したり、動きを統合するためにデザインされた身体のポーズや動き) ディアーナ(マインドフルネス/細心の注意を払うこと) 最初の2つの方法は実践していくうえで、必要不可欠とされている細心の注意を払うことや注意力といった要素が必要となります。注意を払わずに繰り返し行われるポーズは、利点が少ないのに対して、「動きに細心の注意を払い、呼吸に気をつけて行われるものは、全て原則的にはヨガになるのです」(Davis 2009)。 このように細心の注意を払うことや解剖学的精密さにより、ヨガクラスやヨガ療法は以下に付随するテクニックを最大限に用いることになります: 特定のエリアを動かしたり、リラックスさせたり、注意を払ったりするための言葉のキューイングやイメージを用いた導き セラピストによる患者への手技的な調整 変化に気づける固有感覚を促進するための、セラピー前後のボディスキャン セッション間のエクササイズや日常生活動作指導 ハタの限られた枠組みの中で考えてみても、近年のヨガアーサナは目まぐるしいほどのバラエティーに富んでいます。ここでは独断的な立場に立たずに見ていきます;あるヨガが他のものよりも良いとする意図もありません。病院やジム、スパからアスレチックプログラム、村の集会場から修行所といった多種多様な環境において、異なった意図や強度で行われる多くの「ブランド」やバリエーションについても、どれが正しいとすることもありません。 この議論の中では、ヨガの筋膜における効果においてのみ、ヨガを区別していきます: アシュタンガは、高い強度指数を要する力強い形であり、プラクティショナーの心拍数や体内温度をあげます。(Swenson 1999) ヴィンヤサ(フローヨガ)は、形を移行する際にゆっくりとした動作を用いるものです。 ’クラシック’は、肉体的にはより静的なもので(しかし精神的にはダイナミック)、形を正確に保つものです(時に,道具によりサポートされる)。アイアンガヨガやビクラムヨガは、この分野の最たるもの。(Iyengar 1966, Barnett 2003). リストラティブ ー 静的ヨガのバリエーションで、多種多様な完全にサポートされたポーズを用いての、深いリラクゼーションを目的にデザインされたもの(Lasater 1995)。 このリストは全てを網羅しているわけでも、排他的なものでもありません;あるクラスやセラピストはこれらのアプローチを合わせて練習したり、一つのクラスを組み合せて行うこともあります。そしてこれらの他にもヨガの種類は多くあるのです。この著者はアクロヨガを高く掲げ、パートナーヨガにおける対の要素に理解を広げ、ムードラヨガに指をからませ、バクティヨガの信仰的なマントラに身を寄せました、そしてそれはバラエティーに富んだヨガに触れ始めたすぎないのです。 ヨガと筋膜 ヨガが、セラピーとして特定の生理学的コンディションにおいて有益な効果がある、とする研究があります。(Jain et al 1993, Pilkington et al. 2005, Nagendra 1986)。 コントロールした呼吸練習(プラーナヤーマ)の筋膜組織への効果や、生理学的な効果を、他の要素から切り離して計ることは非常に難しいのですが、リサーチは、常識で考えられるポイントをついています。呼吸の増加は、組織により多くの酸素を送り込み、呼吸動作は首から骨盤底に及ぶ胴体を強くし、調和させます。 (Sherman 2005, Kirkwood 2005, Androjna 2008, Farhi 1996, Iyengar 1996). 被験者からは、ヨガやヨガセラピーの後は落ち着き、バランスが取れ、よりエネルギーに溢れていると、事例を交えた報告を受けます。こういった感覚というのは 、 充分に水分が補給されていない組織への水和作用の増加、可動域の増加、以前は固まっていた組織がそれぞれスライドするようになり、受容感覚や神経的統合が高まり、そのエリアが「知覚運動性健忘症」サイクルから目覚める結果からもたらされると想定されます。(Hanna 1998). より明確ではない、生理学的及びスピリチュアル的利点(自律神経系、腺、臓器、心理状態)は、特定のポーズや方法によってあると思われますが、これらの主張を証明することはおろか、役立つコメントを出すことも私たちの能力を超えたものとなります。 メカノバイオロジーにおける挑戦的なリサーチでは、興味深いヨガアーサナの包括的な生理学的影響の可能性を示しています。(Ingber 2003, Langevin 2001, Iatridis 2003, Atance 2004, Arora 1999). 異なった細胞には、異なる力学的な理想環境があることは明らかで、細胞外筋膜基質を経由して届く、インテグリン媒介性のシグナルを感じ取り反応しているのです。 ヨガアーサナと筋筋膜経線 多くのヨガセラピーで共通するアーサナのポーズやストレッチは、単一筋、筋肉グループや結合組織構造に対してのみ働きかけたり、挑戦したりするようにデザインされているのではなく、キネティックチェーン全体や「筋筋膜経線」を連動させるようデザインされています(Myers 2009)。ヨガの正典には千を超えるアーサナやバリエーションがありますが、主なものは「ファミリー」にグループ分けされ、それぞれの経線をより明確にし、異なる部位や問題点が強調されるようにデザインされていると考えられています。(Kraftsow 1999). 参照文献: Atance, J., Yost, M.J., Carver, W. 2004 Influence of the extracellular matrix on the regulation of cardiac fibroblast behavior by mechanical stretch (2004) Journal of Cellular Physiology, 200 (3), pp. 377-386. Androjna C et al,Tissue Engineering Part A. April 2008, 14(4): 559-569. doi:10.1089/tea.2006.0361. Arora, P.D., Narani, N., McCulloch, C.A.G. 1999 The compliance of collagen gels regulates transforming growth factor-β induction of α-smooth muscle actin in fibroblasts (1999) American Journal of Pathology, 154 (3), pp. 871-882. Barnett M 2003 Hot Yoga NY: Barron’s Educational Series Coulter D 2001 Anatomy of Hatha Yoga Honesdale PA: Body and Breath Inc. Davis C 2009 Complementary Therapies in Rehabilitation Thorofare NJ; Slack Inc. Doeser L 2003 The Yoga Directory Edison, NJ: Chartwell Books Farhi D 1996 The Breathing Book NY: Henry Holt & Co Feuerstein G 1998 The Yoga Tradition Prescott AZ: Hohm Press Frederick C & A 2006 Stretch to Win Champaign IL: Human Kinetics Hanna T 1988 Somatics Cambridge MA: Perseus Books Iatrides J et al 2003 Subcutaneous tissue mechanical behavior is linear and elastic under uniaxial tension Connective Tissue Research 44(5): 208-217 Ingber D 2003 Mechanobiology and the diseases of mechanotransduction Annals of Medicine 2003; 33:564-577 Iyengar B 1966 Light on Yoga NY: Allen & Unwin Iyengar B 1996 Light on Pranayama NY: Crossroad Publishing Jain S et al 1993 A study of response pattern of non-insulin dependent diabetics to yoga therapy, Diabetes Res Clin Pract. Jan; 19 (1): 69 - 74 Kaminoff L 2007 Yoga Anatomy Champaign IL: Human Kinetics Kraftsow G 1999 Yoga for Wellness NY: Penguin Arcana Lad V 1984 Aryuveda, A Practical Guide Twin Lakeds WI: Lotus Press Langevin H 2001 et al Mechanical signaling through connective tissue FASEB Journal 2001 15:2275-2282 Lee M 1997 Phoenix Rising Yoga Therapy Deerfield Beach FL: Health Communication Lasater J 1995 Relax and Renew Berkeley CA: Rodmell Press Long R 2005 Scientific Keys, Vol 1 – 3, Bandha Yoga Pub, www.BandhaYoga.com Kirkwood et al 2005 Yoga for anxiety, Brit J of Sports Med 39 (12) pp. 884-891 Mehta S 1990 Yoga The Iyengar Way, NY: Alfred Knopf Mikkonen J et al 2008 A Survey of Musculoskeletal Injury Among Ashtanga Vinyasa Yoga Practitioners, International Journal of Yoga Therapy (18) pp59 – 64 Morse DR et al 1984 A physiological evaluation of the yoga concept of respiratory control of the autonomic bervous system activity; Int. J. Psychsom. 31(1): 3 - 19 Myers T 2009 Anatomy Trains, 2nd ed. Edinburgh: Churchill Livingstone Myers T 1998-99 Kinesthetic Dystonia, J of Bodywork & Movement Therapies 2(2):101-114, 2(4):231-247, 3(1):36-43, 3(2):107-116 Nagendra H et al 1986 An Integrated Approach of Yoga Therapy for Bronchial Asthma Journal of Asthma 22:33, pp 123 – 137 Ornish D 2007 The Spectrum NY: Ballantine Pilkington K et al 2005 Yoga for Depression, the research evidence, J. of Affective Disoiders Dec; 89 (1-3) 13-24, Robin M A 2002 Physiological HandBook for Teachers of Yogasana Tucson AZ: Fenestra Books Seitz D 2010 An Overview of Regulatory Issues for Yoga, Yoga Therapy, and Aryuveda, Int J of Yoga Therapy 2010 (1) pp 1 - 7 Sherman, K.J., et a1.2005. Comparing yoga, exercise, and a self-care book for chronic low back pain: A randomized, controlled trial. Annals of Internal Medicine, 143 (12), 849-56. Smith J et al 2004 Yoga and Pilates London: Hermes House Stiles M Structural Yoga Therapy Boston: Samuel Weiser Books Stiles M 2002 Yoga Sutras of Patanjali Boston: Samuel Weiser Books Stirk J 1988 Structural Fitness London: Elm tree Books Swenson D 1999 Ashtanga Yoga Sugarland TX: Ashtanga Yoga Productions Taylor M 2009 Toga Therapeutics in Davis 2009, Complementary Therapies in Rehabilitation Thorofare NJ; Slack Inc.

ヨガ療法における筋膜 パート2/2
ヨガアーサナと筋筋膜経線 このセクションでは、よくあるポーズやセラピー的動作を、連動されるラインによってグループ分けしました。このプロセスはそれぞれのポーズの複雑さを考慮すると、単純すぎると受け取られるかもしれませんが、ある種の見方やヨガ療法の領域を示すものではあります。 前屈/スーバーフィシャルバックライン スーパーフィシャルバックライン(SBL)は、足指裏から背部を通り頭頂部を渡り眉弓に繋がる一連の筋膜組織ストラップです。SBLの一部や全体への相対的な緊張は、脊柱の一次及び二次湾曲、脚部や足部に影響を及ぼします。すなわちSBLは主要な体重部分を垂直方向に整列させることで、直立バランスを楽に保つための重要な要素となります。 収縮時、SBLの筋肉は身体のほぼ全体を過伸展させます(膝は屈曲しますが)。 伸張時、筋肉と筋膜は胴体と腰部の屈曲に抵抗します。これにより、膝の伸展を保った状態で行われる、身体の屈曲を向上させる様々なストレッチが作り出されます。 図1:スーパーフィシャルバックラインをストレッチするアーサナファミリー、ウッターナサーナ/座位、立位での屈曲などの前屈位、ダウンワードドッグ、ハラアサナ/鍬のポーズ(及びその他関連するショルダースタンド)ボートのポーズ相互的抑制は、SBLとスーパーフィシャルフロントラインの間で起こるために、この姿勢ではディープフロントラインが作用していなければならない。 後屈/スーバーフィシャルフロントライン スーパーフィシャルフロントライン(SFL)は、足指先の表面から太腿の前面、及び胴体部の恥骨から乳様突起までの、身体の表面にある筋筋膜の繋がりを指します。SFLは我々の「柔らかい下腹部」や他の傷つきやすい部分を保護しています。その為、恐怖や保護というパターンにより制限されていることが多くあり、それにより呼吸時の肋骨前部の可動域をせばめることもしばしばあります。 収縮時、SFLは胴体と股関節の屈曲を促しますが、膝は伸展しています。伸張時、SFLは伸展と過伸展に抵抗し、身体全体の動きの中で、この面を開くストレッチを作り出します。 図2:ストレスによりスーパーフィシャルフロントラインが短縮されるため多くのポーズは、このラインの一部分もしくは全体を開くようにデザインされている。シンプルなコブラもしくは「ナマスカーラ/礼拝」の始まりである立位のバックストレッチなどブリッジ徐々に角度を増すバックベンド。スプタ・ヴィラーサナ/仰向けの英雄のポーズ一見簡単に見えるが、短縮した股関節屈曲筋にはチャレンジとなる。 ラテラルライン/側屈 ラテラルライン(LTL)は、足部外側アーチから耳まで、靴紐を通すように交差しながら胴体部分を駆け上がり身体側部にまで及びます。LTLは、日常生活やスポーツにおける体側部の安定性をもたらし、胴体部分を介する小さな歪みを調整します。 収縮時、LTLは同側の側屈と股関節外転を起こします。伸張時には対側の屈曲に抵抗します。LTLを連動させたりストレッチさせるポーズの種類は少なめです。 スパイラルライン/ツイスト スパイラルライン(SPL)は、側頭部から反対側の肩、肋骨、そして背中を通り、同側の腰部ASISに付着する腹斜筋を介して前面部の白線を通るようにして身体の周囲を巡ります。SPLは、腰から太腿前面を通り足根骨の下を通過、下腿側部及び後部を通過して、太腿に渡り、脊柱起立筋から後頭部までをつなぎます。 収縮時、SPLは、どの部分が緊張しているかにより屈曲、伸展、もしくは側屈を行いますが、全体的にSPLは、水平面における捻れやスパイラルの動きを生み出します。伸張時、対側のローテーションに抵抗します。これにより胴体周りや股関節のローテーションによるポーズを作り出します。 図3:捻じりのポーズは、2つのスパイラルラインを連動もしくはストレッチする相互的に鳩のポーズスパイラルライン下部をストレッチ。練習により形がどのように変化していくかが見て取れるー上の写真は経験豊富な教師、中央は生徒、下は初心者。 アームライン/肩と腕のストレッチ アームラインは、筋膜の繋がりであり、中軸骨格から腕の四「面」を介して、指先にまで及ぶ、機能的に繋がった筋筋膜の帯になります。腕の骨周りにおける筋筋膜配列のため、また、腕は安定性よりも可動性に重きを置いて「デザインされている」ため、これらのラインをストレッチするには、多くのポジションやそのライン内でも異なる要素が必要となります。全ての肩の筋肉に対するヨガ療法についてここで述べていくには、ここでの限られた範疇を超えているので、ここでは、一般的な腕のストレッチや強化するポーズが含まれています。 ファンクショナルライン ファンクショナルラインは、片方の上腕骨と反対側の大腿骨を結んだものです ー すなわち、対側の肢帯を結ぶ、とも言えー 身体の前面と背面を通ります。フロント、及びバックファンクショナルライン(FFL、BFL)は、ある意味スパイラルラインのように螺旋状になっているので、多くのスポーツアクションでも見られるような、すべての胴体部の捻りに関わっています。 その為、胴体と股関節屈曲のポーズはBLFをストレッチし、伸展のポーズはFFLをストレッチします。左に胴体部をひねるとFFLの左側とBFLの右側をストレッチし、逆もまた同様です。ここで含まれている2つのポーズは、とりわけファンクショナルラインにチャレンジしています。 ディープロントライン SFLとSBLの間、身体全体にわたって、左右の2本のラテラルラインの間に挟まれ、さらにスパイラル及びファンクショナルラインで包まれ、ディープフロントライン(DFL)は身体の「コア」を形成しています。DFLは、継続した筋筋膜の経路で内側アーチから股下をあがり、鼠蹊部と座骨を通り内腹部組織に繫がります。そして内転筋後部から骨盤底、脊柱前面にあがり、前縦靭帯、腹横筋、横隔膜、縦隔を経て、首から顎及び頭蓋骨下部にまで及びます。 股関節内転と呼吸の機能を除いて、これらのコアマッスルの多くの機能は外側のラインを再現するものとなります。DFLの一部は、胴体もしくは股関節の屈曲を、他の部位は、股関節の伸展を促します。前述のスーパーフィシャルフロントラインの項で述べたバックベンドは、特に生徒が慣れて、よりSFLがストレッチされるようになると、DFLのストレッチとしても用いられるようになります。ダウンワードドッグでは、前述のスーパーフィシャルバックラインでも示されているように、DFL下部の伸びが必要となり、踵を床やマットの上に置いてポーズをとる場合には、しっかりとした足首の背屈ができる必要があります。その為、多くのポーズでは、DFLの要素を連動もしくはストレッチ(またはその両方)することになるのです。 加えて、バランスを取るには全ラインの調和が必要となりますが、とりわけDFL構造の活性化、及び安定性が必要となります。そのため、この項では、DFLとその周辺ライン両方の活性化が必要となる、一般的なバランスのポーズを取り入れています。 図4:ディープフロントライン連動させることがヨガとピラティスのゴールとなるが、多くのビギナーの生徒は「コア」の筋肉を正しく使うことができず、それらが全ての姿勢で使われる事は難しい。バランスのポーズ逆転のポーズプランクのポーズのようにコアの筋肉の動員が必要。山のポーズのような「自己想起」の練習サヴァーサナディープフロントラインを連動させることも重要。 ヨガ療法トレーニングのスタンダード 買い手の危険負担原則。西ヨーロッパとアメリカ北部ではー実際には全世界で(インドのヨガ教師総数よりも、カリフォルニアにいるヨガ教師数の方が多いーDavis 2009)ー 職業としてのヨガ教育とヨガ療法は、早期発達を遂げました。そのため、トレーニングのスタンダード及び専門的能力の保証というものは、未だ初期段階にあり、スキルレベルにも幅があります。多くの場合、200時間のトレーニングを経て有資格のヨガ教師となりますが、その基準を500時間(もしくはそれ以上)にし、多くのヨガセラピストが要するトレーニングとすることが計画されています(Seitz 2010)。 ヨガセラピストや教師を探す際には、そのセラピスト/教師が国際ヨガセラピスト協会(IAYT)か、ヨガアライアンス(YA)に属しているか、もしくは何かしらの資格を持っているかを調べることができます。(www.iayt.org, www.yogaalliance.org)ヨガセラピストの中には、理学療法士、作業療法士、もしくは公認看護士であったり、アスリートのコーチやピラティス、パーソナルトレーニングに従事している人もいます。 ヨガにおける怪我は珍しくはないものの、著者の経験及び進行中のリサーチは、それらの怪我は、高圧的なセラピストや下手な導き方によって起こってしまうというよりは、熱心な生徒が無理にオーバーストレッチさせて怪我につながってしまうケースが多いようです(Mikkonen et al 2008)。 参照 ヨガ療法は、いわば二千年にも及ぶ「現場研修」のようなもので、近年の臨床的な実験や、繰り返し行われるエビデンスに基づいたリサーチというのは、この分野ではまだ始まったばかりです。また、ヨガセラピーには心理学的及び有機生物学的な利点があるというのも、事例証拠を元に示すことができると思いますが、これらはまだ正式な研究により充分にサポートされているわけではありません(Ornish 2007, Morse et al 1984)。 筋膜リリーステクニックとして治療的に使われるヨガは、理学療法士やいわゆる「代替」療法といった他の多くの徒手療法と似ているところが多くあります。ヨガの利点というのは、しばしばパーソナルトレーニングや理学療法で行われている、単なる局所的な強化や、徒手療法で用いられる受動的な組織のリリーステクニックとは一線を画し、ストレッチ、強化、バランスのコンビネーションから産まれるのです。 セラピーとしてのヨガは、これらの関連するポーズの「家族写真」から見て取れるように、明らかに身体全体を走る運動学上の、そして筋膜の鎖を調整し繋げていきます。才能溢れるセラピストや教師の手にかかれば、オープンで強くバランスの取れた身体へと導かれることでしょう。セラピストの中には、高齢者を専門としている人もいれば、産前、産後、子供向け、身体の自由度が異なる方やその他の対象グループを専門としている人もいます。レストレーティブヨガのポーズは身体の不自由な方にもよく使われていますが、多くのヨガクラスやプライベートセラピーセッションにおいては、ポーズを取ることが身体的に可能で、意欲的な人が対象となります。 参照文献: Atance, J., Yost, M.J., Carver, W. 2004 Influence of the extracellular matrix on the regulation of cardiac fibroblast behavior by mechanical stretch (2004) Journal of Cellular Physiology, 200 (3), pp. 377-386. Androjna C et al,Tissue Engineering Part A. April 2008, 14(4): 559-569. doi:10.1089/tea.2006.0361. Arora, P.D., Narani, N., McCulloch, C.A.G. 1999 The compliance of collagen gels regulates transforming growth factor-β induction of α-smooth muscle actin in fibroblasts (1999) American Journal of Pathology, 154 (3), pp. 871-882. Barnett M 2003 Hot Yoga NY: Barron’s Educational Series Coulter D 2001 Anatomy of Hatha Yoga Honesdale PA: Body and Breath Inc. Davis C 2009 Complementary Therapies in Rehabilitation Thorofare NJ; Slack Inc. Doeser L 2003 The Yoga Directory Edison, NJ: Chartwell Books Farhi D 1996 The Breathing Book NY: Henry Holt & Co Feuerstein G 1998 The Yoga Tradition Prescott AZ: Hohm Press Frederick C & A 2006 Stretch to Win Champaign IL: Human Kinetics Hanna T 1988 Somatics Cambridge MA: Perseus Books Iatrides J et al 2003 Subcutaneous tissue mechanical behavior is linear and elastic under uniaxial tension Connective Tissue Research 44(5): 208-217 Ingber D 2003 Mechanobiology and the diseases of mechanotransduction Annals of Medicine 2003; 33:564-577 Iyengar B 1966 Light on Yoga NY: Allen & Unwin Iyengar B 1996 Light on Pranayama NY: Crossroad Publishing Jain S et al 1993 A study of response pattern of non-insulin dependent diabetics to yoga therapy, Diabetes Res Clin Pract. Jan; 19 (1): 69 - 74 Kaminoff L 2007 Yoga Anatomy Champaign IL: Human Kinetics Kraftsow G 1999 Yoga for Wellness NY: Penguin Arcana Lad V 1984 Aryuveda, A Practical Guide Twin Lakeds WI: Lotus Press Langevin H 2001 et al Mechanical signaling through connective tissue FASEB Journal 2001 15:2275-2282 Lee M 1997 Phoenix Rising Yoga Therapy Deerfield Beach FL: Health Communication Lasater J 1995 Relax and Renew Berkeley CA: Rodmell Press Long R 2005 Scientific Keys, Vol 1 – 3, Bandha Yoga Pub, www.BandhaYoga.com Kirkwood et al 2005 Yoga for anxiety, Brit J of Sports Med 39 (12) pp. 884-891 Mehta S 1990 Yoga The Iyengar Way, NY: Alfred Knopf Mikkonen J et al 2008 A Survey of Musculoskeletal Injury Among Ashtanga Vinyasa Yoga Practitioners, International Journal of Yoga Therapy (18) pp59 – 64 Morse DR et al 1984 A physiological evaluation of the yoga concept of respiratory control of the autonomic bervous system activity; Int. J. Psychsom. 31(1): 3 - 19 Myers T 2009 Anatomy Trains, 2nd ed. Edinburgh: Churchill Livingstone Myers T 1998-99 Kinesthetic Dystonia, J of Bodywork & Movement Therapies 2(2):101-114, 2(4):231-247, 3(1):36-43, 3(2):107-116 Nagendra H et al 1986 An Integrated Approach of Yoga Therapy for Bronchial Asthma Journal of Asthma 22:33, pp 123 – 137 Ornish D 2007 The Spectrum NY: Ballantine Pilkington K et al 2005 Yoga for Depression, the research evidence, J. of Affective Disoiders Dec; 89 (1-3) 13-24, Robin M A 2002 Physiological HandBook for Teachers of Yogasana Tucson AZ: Fenestra Books Seitz D 2010 An Overview of Regulatory Issues for Yoga, Yoga Therapy, and Aryuveda, Int J of Yoga Therapy 2010 (1) pp 1 - 7 Sherman, K.J., et a1.2005. Comparing yoga, exercise, and a self-care book for chronic low back pain: A randomized, controlled trial. Annals of Internal Medicine, 143 (12), 849-56. Smith J et al 2004 Yoga and Pilates London: Hermes House Stiles M Structural Yoga Therapy Boston: Samuel Weiser Books Stiles M 2002 Yoga Sutras of Patanjali Boston: Samuel Weiser Books Stirk J 1988 Structural Fitness London: Elm tree Books Swenson D 1999 Ashtanga Yoga Sugarland TX: Ashtanga Yoga Productions Taylor M 2009 Toga Therapeutics in Davis 2009, Complementary Therapies in Rehabilitation Thorofare NJ; Slack Inc.
筋膜フィットネス:神経筋筋膜ウェブのトレーニング パート1/3
リサーチは、エクササイズや動きに関わる脳に、今までとは異なったアプローチを取ることが、未来への新しい波である、その理由を示しています。 フィットネストレーニングにおける筋膜の役割に興味を持っているのであれば、以下の質問は新しい扉を開くことになるでしょう。 多くの怪我は結合組織(筋膜)の損傷であり、筋肉の損傷ではありませんー それではどのようにして怪我を予防、ダメージを修復し、伸縮性や弾力性をシステムに築き上げていくのが最善の方法なのでしょうか。 筋膜には筋肉の10倍もの感覚神経終末があります;どのようにして筋膜と筋肉に固有受容刺激を与えることができるのでしょうか? 伝統的な筋肉や筋膜の解剖学テキストは、動作機能における基本的な理解不足という点で正確ではありません。それでは我々はどのように「安定の臓器システム」である筋膜を全体として捉えて、働きかけることができるでしょうか? 意識的にしろ、無意識的にしろ、あなたは動きに関わるキャリア全体において、筋膜に対して働きかけていましたーそれは避けることができないものなのです。しかしながら、新しいリサーチは、ファンクショナルトレーニングにおける筋膜とその他結合組織の重要性を強調しています(筋膜学術講演会 2009)。筋膜は「筋肉の周りにあるプラスチック製のラップ」というだけではないのです。「筋膜は安定性と物理的な力の伝達に作用する臓器システムなのです(Varela & Frenk 1987)。 これを理解することは「フィットネス」に関する私達の考え方を革新することになるかもしれません。筋膜ネットに関するリサーチにより、伝統的な信念や近年好まれているものが揺らぐことになります。これらのエビデンスは人生におけるフィットネスすべてへの新たな考察に向けられていますーですから筋膜フィットネスという表現なのです。 この記事では、全体像としての筋膜ネットのイメージを浮かび上がらせ、及び筋膜ネットを鍛える為の理解をより深めることができる、最近のリサーチの数多くの側面の中から3点について取り上げ、探求していきます。 神経筋筋膜のウェブ 筋膜は身体組織のシンデレラであるーシステム的には無視をされ、解剖では細かく切り裂かれ捨てられる(Schleip 2003)。しかしながら、筋膜は生理学的容器であり、すべての臓器(筋肉を含む)にとってのコネクターなのである。解剖において、筋膜は文字通り油ぎってぐちゃぐちゃしたものである(教科書に描かれているものとは似ても似つかないもの)、そして各個人により千差万別で、実際の様子を描くことは非常に難しい。多くの理由から、筋膜はシステム全体として捉えられていませんでした;そのため私たちは、筋膜の生体力学における全体の役割について無知であったのです。 ありがたいことに、統合された機械生物学的筋膜ウェブの性質は、よりはっきりとしてきました。それはまさに頭頂からつま先まで、皮膚からコアまで、そして生誕から死まで、分かれ目のない一つのネットなのです(Shultz & Feitis 1996)。体内のすべての細胞は筋膜の張力環境に繋がってーそして反応してーいます(Ingber 1998)。メカニクスを変化させると、細胞もその機能を変化させます(Horwitz 1997)。“空間医療”の一部として、ストレッチ、強化、そして形を変えていくことーこれはパーソナルトレーニングの革新的で新しい捉え方ですー(Myers 1998)。 事実を基にすれば、多くの人は、筋膜を軽視する筋骨格系よりも神経筋筋膜という言葉の方を好むでしょう。(Schleip 2003)。足底筋膜、アキレス腱、腸脛靭帯、胸腰腱膜、項靭帯等、私達は、筋膜ウェブの中に存在する個々の構造を識別することに馴染んでいますが、これらは単に一つの筋膜ウェブにおけるエリアにつけられた便利なラベルにしかすぎないのです。それは郵便番号のようなものと言っても差し支えがないかもしれませんが、それぞれが分離した構造ではないのです。(サイドバーの「筋肉の孤立化」vs「筋膜の統合」をご覧ださい)。 大西洋、太平洋、もしくは地中海について話をすることはできます、でも海は繋がっていて世界にたった一つしかないのです。筋膜も同様です。私達は、個々の神経についての話をしますが、神経系統は全体として反応することを知っています。筋膜のウェブはどのようにしてシステムとして機能するのでしょうか? 魔法のように全体として筋膜のウェブを取り出したとしたら、それは裏も表も身体のすべての型を見せてくれることになります。それは大きな一つのネットで、その中で筋肉は魚のようにうごめいているのです。臓器はクラゲのようにぶら下がっています。全てのシステム、すべての臓器、すべての細胞までも、単一の筋膜ネットの海の中に埋め込まれているのです。 このコンセプトは重要です。なぜなら私たちは、個々の構造に名称を与え、臨床的にもそのように間挙げる傾向が強くあります:「上腕二頭筋を断裂してしまったね」、というように「上腕二頭筋」が我々のコンセプトであることを忘れてしまっています。私達が共通に使う、科学的専門用語というのは間違った印象を与えます。それにたいしてニューエイジの合言葉は文字通りに、真実を告げています;身体 (特に筋膜ネット)は、ひとつの繋がったまとまりで、その中で筋肉と骨が浮かんでいるのです。 怪我によりこのネットが破れたり、外科医のメスで切られたり、栄養や水分補給を充分に行うことができたり、もしくは果糖がたっぷりはいったコーンシロップで、詰まらせてしまったりします。どのように筋膜を扱ったとしても、いつかはその弾力性を失っていきます。例えば目のレンズにおいては、そのネットは通常固くなり、およそ50歳で老眼用のメガネが必要になるというようなことです。 皮膚においては、ネットの擦り切れがシワになります。股関節の軟骨のような、とても重要な要素も、命が途絶えるまでもたずに修繕が必要となるかもしれません。しかしながら最後の息を引き取るその瞬間まで、筋膜のウェブは生まれた時と同じ一枚のネットなのです。 このシステムが,神経系や循環系のように複雑なシグナルやホメオスタシスのメカニクスを発達させるというのは、特に驚くべきことでもありません(Langevin et al. 2006)。私たちが、この結合組織システムの反応を今まで見たり、探求したりしてこなかったことは大きな驚きであります。 筋肉の孤立化 vs 筋膜の統合 フィットネスの専門家の多くは、孤立化した筋肉の機能を学んできました。基本的に西洋キネシオロジーにおける解剖学は、このように問いかけます。:「上腕二頭筋が骨格の中で唯一の筋肉だとしたら、それはどのようなアクションを起こしますか?」 上腕二頭筋のみを取り上げれば、この筋肉は橈骨尺骨の回外筋であり、肘の屈曲筋であり、多少肩を斜めに屈曲させる筋肉でもあります。こう言ったことを覚えた時、私たちは上腕二頭筋がどういうものでそれが何をするものかを理解した気になります。それはひとつのものの見方ではあります。 ただ、上腕二頭筋というのは決して単独で働いているわけではないのです。その機能を知るために、筋肉を孤立化していくというのは統合(インテグレーション)や全体性(ホーリズム)とは正反対のものになります。実践的な違いは、どのようなものでしょうか?筋肉のみを単独で学ぶことは、日々の筋肉の機能において重要な、4つの筋膜の要素を取り残してしまうことになります。 隣接する内側及び外側に位置する筋肉からの影響と、それらの筋肉への影響。上腕二頭筋は烏口上腕筋、上腕筋、そして回内筋、さらには上腕三頭筋の隔膜にまで力を伝達する筋膜的な繋がりを持ちます。これらの筋膜的な繋がりは、上腕二頭筋や腕の機能に影響を及ぼします(Huijing 2007)。 近位及び遠位で繋がっている筋肉からの影響と、それらの筋肉への影響。上腕二頭筋は骨間膜、橈骨周辺の筋膜、及び屈曲筋に入り込む上腕二頭筋腱膜と遠位で繋がっています。近位では短頭と長頭を介して、それぞれ小胸筋と棘上筋と繋がっています。(Myers 2001, 2009)。 筋収縮がそのエリアの靭帯に及ぼす影響。上腕二頭筋を収縮させることは、肩と肘、両方の靭帯を安定させます。私たちの靭帯が筋肉と平行に位置しているという憶測は正しいものではありません。多くの靭帯は、連続して、ダイナミックに筋肉と力学的に統合されているため、筋収縮は靭帯が全ての角度で関節を安定させる助けとなるのです(Van der Wal 2009)。 全ての筋肉は、神経や血管により供給されなければならないという事実。これらの「ワイヤーやチューブ」は筋膜のシーツに包まれて届けられます。これらのシーツが悪い姿勢により捻れていたり、詰まっていたり、短くなりすぎている場合、筋肉の機能に影響を及ぼします(Shacklock 2005)。 (パート2/3はこちらへ) 参考文献 Chen, C.S., et al. 1997. Geometric control of cell life and death. Science, 276 (5317), 1425–28. Chino, K., et al. 2008. In vivo fascicle behaviour of synergistic muscles on concentric and eccentric plantar flexion in humans. Journal of Electromyography and Kinesiology, 18 (1), 79–88. Desmouliere, A., Chapponier, C., & Gabbiani, G. 2005. Tissue repair, contraction, and the myofibroblast. Wound Repair Regeneration, 13 (1), 7–12. Fascia Congress. 2009. www.fasciacongress.org/2009. Fuller, R.B. 1975. Synergetics. New York: Macmillan. Grinnell, F. 2008. Fibroblast mechanics in three-dimensional collagen matrices. Trends in Cell Biology, 12 (3), 191–93. Grinnell, F., & Petroll, W. 2010. Cell motility and mechanics in three-dimensional collagen matrices. Annual Review of Cell and Developmental Biology, 26, 335–61. Horwitz, A. 1997. Integrins and health. Scientific American, 276, 68–75. Huijing, P. 2007. Epimuscular myofascial force transmission between antagonistic and synergistic muscles can explain movement limitation in spastic paresis. Journal of Biomechanics, 17 (6), 708–24. Huijing, P.A., & Langevin, H. 2009. Communicating about fascia: History, pitfalls and recommendations. In P.A. Huijing et al. (Eds.), Fascia Research II: Basic Science and Implications for Conventional and Complementary Health Care. Munich, Germany: Elsevier GmbH. Iatrides, J., et al. 2003. Subcutaneous tissue mechanical behaviour is linear and viscoelastic under uniaxial tension. Connective Tissue Research, 44 (5), 208–17. Ingber, D. 1998. The architecture of life. Scientific American, 278, 48–57. Ingber, D. 2008. Tensegrity and mechanotransduction. Journal of Bodywork and Movement Therapies, 12 (3), 198–200. Kubo, K., et al. 2006. Effects of series elasticity on the human knee extension torque-angle relationship in vivo. Re- search Quarterly for Exercise & Sport, 77 (4), 408–16. Langevin, H. 2006. Connective tissue: A body-wide signaling network? Medical Hypotheses, 66 (6), 1074–77. Langevin, H., et al. 2010. Fibroblast cytoskeletal remodeling contributes to connective tissue tension. Journal of Cellular Physiology. E-pub ahead of publication. Oct. 13, 2010. Myers, T.W. 1998. Kinesthetic dystonia. Journal of Bodywork and Movement Therapies, 2 (2), 101–14. Myers, T.W. 2009. Anatomy Trains: Myofascial Meridans for Manual and Movement Therapists. New York: Churchill- Livingston. Sbriccoli, P., et al. 2005. Neuromuscular response to cyclic loading of the anterior cruciate ligament. The Amercian Journal of Sports Medicine, 33 (4), 543–51. Schleip, R. 2003. Fascial plasticity—a new neurobiological explanation. Journal of Bodywork and Movement Therapies. Part 1: 2003, 7 (1), 11–19; part 2: 2003, (2), 104–16. Shacklock, M. 2005. Clinical Neurodynamics. Burlington, MA: Butterworth-Heinemann. Shultz, L., & Feitis, R. 1996. The Endless Web. Berkeley, CA: North Atlantic Books. Snyder, G. 1975. Fasciae: Applied anatomy and physiology. Kirksville, MO: Kirksville College of Osteopathy. Stecco, C., et al. 2009. Treatment of phantom-limb pain according to the fascial manipulation technique: A pilot study. In P.A. Huijing et al. (Eds.), Fascia Research II: Basic Science and Implications for Conventional and Complemen- tary Health Care. Munich, Germany: Elsevier GmbH. Stillwell, D.L. 1957. Regional variations in the innervation of deep fasciae and aponeuroses. The Anatomical Record, 127 (4), 635–53. Taguchi, T., et al. 2009. The thoracolumbar fascia as a source of low back pain. In P.A. Huijing et al. (Eds.), Fascia Re- search II: Basic Science and Implications for Conventional and Complementary Health Care. Munich, Germany: El- sevier GmbH. Van der Wal, J. 2009 The architecture of the connective tissue in the musculoskeletal system: An often overlooked functional parameter as to proprioception in the locomotor apparatus. In P.A. Huijing et al. (Eds), Fascia Research II: Basic Science and Implications for Conventional and Complementary Health Care. Munich, Germany: Elsevier GmbH. Varela, F., & Frenk, S. 1987. The organ of form. Journal of Social and Biological Structures, 10 (1), 1073–83. Yahia, L.H., Pigeon, P., & DesRosiers, E.A. 1993. Viscoelastic properties of the human lumbodorsal fascia. Journal of Biomedical Engineering 15, 425–29.
筋膜フィットネス:神経筋筋膜ウェブのトレーニング パート2/3
言語の定義 医療において筋膜という言葉は、腱、靭帯、その他の組織と区別するために特定の形態 と組織学を持つものに対して用いられます。しかしながら、この記事においては他に一般化された言葉がないため、この系統だてられた結合組織ネット全体を指す名称として筋膜という表現を使用しています(Huijing & Langevin 2009)。 結合組織には、血管、血腋細胞その他の、私たちが検証しようとしている構造ネットの一部ではない要素も含まれます。おそらく、もっとも近い単語は細胞外基質(ECM)となるでしょう。ECMには細胞質ではない体内全てのものが含まれます。ECMには3つの主な要素が含まれます。 繊維:強くしなやかな織組織。主にコラーゲンで成りたっており(12種類に及ぶ)、その親族としてエラスチンやレチクリンがある ー 共に、コンパートメントを分け、お互いを結びつける。 糊:ヘパリン、フィブロネクチン、ヒアルロン酸のように変化しやすくコロイド状のゲルで、変化に順応し、神経や上皮といったその他の細胞に基質を提供する。 水分:細胞の周囲に存在し、交換手段として浸透する液体;糊と混ざり合い異なった特性を持つ物質を作り出す;また繊維を保湿し、しなやかさを保つ役割も担う。 筋膜形態のほんの一例 この記事では、一般化された筋膜という単語を相互に繋がっている繊維と糊の網を表すために使用しています。A:2つの筋肉は「けばだったもの」でまとめられている ー 疎性組織。B:「縛り付けるテープ」大腿四頭筋を包む筋膜の性質。C:(J-C Guimbertau博士の快諾により掲載)我々の皮膚の下、筋肉の間、そしてお互いをスライドしなければならない解剖学組織の、どの場所においても変化と動きを許容する非常に繊細で粘着質な組織。 以下のトピックでECMについて扱いますが、私達が定義する筋膜という言葉は繊維と糊を増加させ、怪我、トレーニング、及び習慣による要求に対する反応として、組織のリモデルを行う繊維芽細胞やマスト細胞を含みます。 ECMを構成する主な構造的要素は、コラーゲン繊維、エラスチン、そしてレチクリンです。コラーゲンはこの中でも最も一般的で、圧倒的な強度を誇ります。これは肉にみられる、白くて丈夫なものです。コラーゲン繊維は三重螺旋になっており、もしも0.5インチ(約1、27cm)の厚みがあるとすれば、約1ヤード(0.91m)の長さになり、昔の三つ編みロープのように見えます(Snyder 1975)。コラーゲン繊維は、腱や靭帯のように規則的な配列で配置されることもあれば(規則的密集)、フェルトのようにランダムに交差することもあります(不規則的密集、もしくは不規則的散在)。 コラーゲン繊維は、実際に相互にはり付くことはできず、ムコ多糖体であるグリコアミノグリカン(GAGs)と呼ばれるたんぱく質(どちらも鼻水を表す長い単語)によって結合されます。私たちは、その化学物質を多少変化させることによって、驚くべき特性の配列を示し、分厚く粘着的なものから水々しい潤滑油となる粘液やコロイド状物質によって、まとめられています。 シダの葉状の粘液分子(これは親水性のものです)は水分を吸収するために開いたり、水分が存在しない時には閉じてお互いを結びつけます。この粘液分子は化学物質によって、各層を結びつけるか、もしくは各々がスライドするように働きかけます(Grinnell 2008)。 私たちが「ストレッチ」と呼ぶ現象や伸長すること(科学者は「クリープ / 漸動」と呼んだりヒステレシスと呼んだりします)は、コラーゲン繊維が伸張しているのではなく、水分を含んだGAGsの糊の上を繊維がお互いにスライドしているのです(Sbriccoli et al. 2005)。GAGsから水気が取り除かれると、組織は非常にストレッチしにくくなります(Schleip 2003)。 怪我の多くは、結合組織が反応できるよりも早くストレッチされてしまうことで起こります。水分が少なければ少ないほど、伸縮的反応は失われます。 身体の電気? 結合組織細胞は、繊維とGAGsを生産します。そしてそれらの素材は驚く程多くのバラエティー豊富な建築素材に形をかえます。もしも身体構造をHome Depot®(アメリカ最大のホームセンターチェーン)で買えるもので作り変えるとしたら、いったい何が必要でしょうか? 骨には木材、もしくは塩ビ菅、軟骨組織にはシリコンゴム、大量の紐、ワイヤー、チューブ、プラスチック製のシート、ゴムバンド、綿、ネット、油脂にオイル − リストは延々と続きます。ガムテープなしで身体を作ろうとしますか? 身体は、ECMの繊維と糊を様々な配合比で混ぜ、異なった方法で化学変化を起こし、これら素材すべてを製造し、さらに多くの素材を作り出します(Snyder 1975)。骨には繊維基質がありますが −革のようなものです−粘液的な基質が ミネラル塩と系統的に入れ替えられています。 軟骨には、同じように革のような基材が存在しますが、糊は 繊維質の革に浸透する、丈夫でありながらしなやかな“プラスチック”の中で乾いてしまっています。靭帯や腱では、ほとんどの糊が絞り出されています。血液や関節液では、繊維は空気に触れるまでは液体状で存在するのみで、空気に触れるとかさぶたになります。 身体にあるこの工場は魅惑的です:歯の象牙質、歯茎、心臓弁、そして透明な目の角膜でさえも、このようにして作られているのです。 ディープ・フロントアーム・ライン アナトミートレインではひとつの筋肉を繋げ合う、筋膜的繋がりを示しています – パート1/3で示す孤立した上腕二頭筋から−、機能する全体へと。 再構築とテンセグリティー トレーニングの観点では、筋肉が身体を形作るかもしれませんが、結合組織は全体性として、私達の身体の形を作ります。結合組織は骨を結びつけ、押し出す骨を引き寄せます(テンセグリティーシステムのように) ECMは様々な方法で、自らを再構築することができます(Chen et al. 1997)。それは筋肉がトレーニングに反応して再構築するように、細胞からの直接的なシグナル (Langevin et al. 2010); 怪我 (Desmouli`ere, Chapponnier & Gabbiani 2005); 長期間に渡る力学的な力 (Iatrides et al. 2003); 利用パターン(感情的なものを含む);重力;そして体内で起こる特定の化学反応によって、筋膜は自らを再構築します (Grinnell & Petroll 2010)。 再構築の複雑性については、研究室で調査され始めたばかりです;これからの10年で詳細が明らかにされることでしょう。 テンセグリティーのアイデア(張力と統合)と再構築の現象は、ヨガやロルフィング®、やストラクチュラルインテグレーションとして知られるタイプのマニュアルセラピー、そしてフォームローリングを含む深部組織関連等の構造セラピーの基礎となります。 要求を変えることで −ボディーワークやパーソナルトレーニングで行うように− 筋膜システムも、その新たな要求に対して反応をします。この共通したテーマはマニュアルセラピーとムーブメントトレーニングが組合わさり 姿勢や機能を自然な状態に修正すること; 後々大きな問題にならないように小さな問題に対処すること; 長期間の怪我によってもたらされる結果を和らげること;そして ファンクショナルムーブメントを年齢尺度のより上のほうまで広げること に対してパワフルな手法となる未来を指し示しています 。 (パート3/3はこちらへ) 参考文献 Chen, C.S., et al. 1997. Geometric control of cell life and death. Science, 276 (5317), 1425–28. Chino, K., et al. 2008. In vivo fascicle behaviour of synergistic muscles on concentric and eccentric plantar flexion in humans. Journal of Electromyography and Kinesiology, 18 (1), 79–88. Desmouliere, A., Chapponier, C., & Gabbiani, G. 2005. Tissue repair, contraction, and the myofibroblast. Wound Repair Regeneration, 13 (1), 7–12. Fascia Congress. 2009. www.fasciacongress.org/2009. Fuller, R.B. 1975. Synergetics. New York: Macmillan. Grinnell, F. 2008. Fibroblast mechanics in three-dimensional collagen matrices. Trends in Cell Biology, 12 (3), 191–93. Grinnell, F., & Petroll, W. 2010. Cell motility and mechanics in three-dimensional collagen matrices. Annual Review of Cell and Developmental Biology, 26, 335–61. Horwitz, A. 1997. Integrins and health. Scientific American, 276, 68–75. Huijing, P. 2007. Epimuscular myofascial force transmission between antagonistic and synergistic muscles can explain movement limitation in spastic paresis. Journal of Biomechanics, 17 (6), 708–24. Huijing, P.A., & Langevin, H. 2009. Communicating about fascia: History, pitfalls and recommendations. In P.A. Huijing et al. (Eds.), Fascia Research II: Basic Science and Implications for Conventional and Complementary Health Care. Munich, Germany: Elsevier GmbH. Iatrides, J., et al. 2003. Subcutaneous tissue mechanical behaviour is linear and viscoelastic under uniaxial tension. Connective Tissue Research, 44 (5), 208–17. Ingber, D. 1998. The architecture of life. Scientific American, 278, 48–57. Ingber, D. 2008. Tensegrity and mechanotransduction. Journal of Bodywork and Movement Therapies, 12 (3), 198–200. Kubo, K., et al. 2006. Effects of series elasticity on the human knee extension torque-angle relationship in vivo. Re- search Quarterly for Exercise & Sport, 77 (4), 408–16. Langevin, H. 2006. Connective tissue: A body-wide signaling network? Medical Hypotheses, 66 (6), 1074–77. Langevin, H., et al. 2010. Fibroblast cytoskeletal remodeling contributes to connective tissue tension. Journal of Cellular Physiology. E-pub ahead of publication. Oct. 13, 2010. Myers, T.W. 1998. Kinesthetic dystonia. Journal of Bodywork and Movement Therapies, 2 (2), 101–14. Myers, T.W. 2009. Anatomy Trains: Myofascial Meridans for Manual and Movement Therapists. New York: Churchill- Livingston. Sbriccoli, P., et al. 2005. Neuromuscular response to cyclic loading of the anterior cruciate ligament. The Amercian Journal of Sports Medicine, 33 (4), 543–51. Schleip, R. 2003. Fascial plasticity—a new neurobiological explanation. Journal of Bodywork and Movement Therapies. Part 1: 2003, 7 (1), 11–19; part 2: 2003, (2), 104–16. Shacklock, M. 2005. Clinical Neurodynamics. Burlington, MA: Butterworth-Heinemann. Shultz, L., & Feitis, R. 1996. The Endless Web. Berkeley, CA: North Atlantic Books. Snyder, G. 1975. Fasciae: Applied anatomy and physiology. Kirksville, MO: Kirksville College of Osteopathy. Stecco, C., et al. 2009. Treatment of phantom-limb pain according to the fascial manipulation technique: A pilot study. In P.A. Huijing et al. (Eds.), Fascia Research II: Basic Science and Implications for Conventional and Complemen- tary Health Care. Munich, Germany: Elsevier GmbH. Stillwell, D.L. 1957. Regional variations in the innervation of deep fasciae and aponeuroses. The Anatomical Record, 127 (4), 635–53. Taguchi, T., et al. 2009. The thoracolumbar fascia as a source of low back pain. In P.A. Huijing et al. (Eds.), Fascia Re- search II: Basic Science and Implications for Conventional and Complementary Health Care. Munich, Germany: El- sevier GmbH. Van der Wal, J. 2009 The architecture of the connective tissue in the musculoskeletal system: An often overlooked functional parameter as to proprioception in the locomotor apparatus. In P.A. Huijing et al. (Eds), Fascia Research II: Basic Science and Implications for Conventional and Complementary Health Care. Munich, Germany: Elsevier GmbH. Varela, F., & Frenk, S. 1987. The organ of form. Journal of Social and Biological Structures, 10 (1), 1073–83. Yahia, L.H., Pigeon, P., & DesRosiers, E.A. 1993. Viscoelastic properties of the human lumbodorsal fascia. Journal of Biomedical Engineering 15, 425–29.

筋膜フィットネス:神経筋筋膜ウェブのトレーニング パート3/3
どのように神経筋筋膜ウェブを鍛えるか もしも筋膜が、唯一の空間を組織する調整可能なテンセグリティーで、身体全体に行き届き(局所的にも全体的にも)張力や圧縮の生体力学を調整しているのであれば、このように問うことができます:「筋肉や神経制御への働きかけとともに、怪我を予防、修復し、システムに弾性をもたらす為には、どのようにこのシステムを鍛えることができるのだろうか?」 この質問に対する答えはまだ発展途上ですが、実験室とトレーニングの現場両方において急速に展開されています。発展を遂げ、伝統的に使用されている私たちがイメージするものや実践していることを裏付けるリサーチもあります。 ここでは、私たちのアイデアを変える(もしくはもうすぐに)神経筋筋膜ウェブが実際にどのようにして働き、結合組織が人生におけるフィットネス全体を発展させるのにどのように作用するのかといった、いくつかの驚くべき発見に焦点をあてます 。これらの更に詳細な結果は www.fascialfitness.de や www.anatomytrains.com の筋膜フィットネスのセクションで見つけることができます。 発見#1:特定のトレーニングは全身の弾性エネルギーに必要不可欠な筋膜弾性を向上させることができる。 筋膜弾性は最近まで認識されておらず、それに関わるメカニズムも現在研究されている段階です (Chino et al. 2008)。にもかかわらずトレーニングに適用されていることは既に明らかです。基本的なニュースは、結合組織は − 腱や腱膜といった密生組織でさえも− 以前考えられていたよりも更に、かなりの弾力性があるということです。 ニュースの重要な2つ目の部分は、筋膜弾性はとても素早く蓄積され返還されます。言い換えれば、ナーフボール(Nerf™ ball/スポンジ製のボール)というよりはスーパーボールのような感じなのです。そのため、筋膜弾性はランニングやウォーキング、または細かいジャンプというように循環し素早く繰り返される動きの時にのみ関わる要因となりますが、自転車漕ぎは含まれません、なぜなら筋膜弾性の特質を利用するには、反復サイクルが遅すぎるからです。 ランニング時におけるふくらはぎの伸長度の計測は、筋肉が等尺性収縮をしている際に、筋膜の弾力性伸縮により、背屈に必要とされる長さがもたらされることを示しました (Kubo et al. 2006)。 これは、腱には弾力性がなく、足が地面に着く前後に起こる循環動作の間、筋肉が伸び縮みすると、私たちが以前理解していたこととは矛盾します。 トレーニングを積み、この弾力性を利用するランナーは、伸張時にエネルギーを蓄え、リリースの時に返還することで、より少ない筋力をランニング時に使用しています。(「より少ないグルコース」:参照)そのため、彼らは、より長い距離を少ない疲労で走ることができるのです。 この弾力性を構築するには、組織にそのように作動するよう働きかけることです。これをゆっくりと行うこと(筋力トレーニングと比べて)は筋膜トレーニングにおいては絶対的な特性となります(筋膜弾性を構築するには6−24ヶ月かかります)。 必要なこと: バウンシング(反発性)。母指球で着地する時、身体は減速や加速を行うことに弾力性を利用しているだけでなく、腱や筋膜システム全体に弾力性を構築しています。最善のトレーニング効果は快楽原理に基づくようです:優雅さと、最小の努力と最大のたやすさの理想的な共鳴を感じるのです。 準備としての対抗運動。対抗運動をして動作の準備をすることは(例えば、伸展して立ち上がる前に屈む、投球前のワインドアップ、またはケトルベルを身体から離す前に身体に近づける) 動作の円滑化を助けるために、筋膜弾性のパワーを最大限に利用することになります。 必要ではないこと: ぎくしゃくした動きと突然の方向転換。縄跳びで踵だけでの着地をすることを想像してみてください。この時、システム全体にかかるストレスは甚大で、筋膜システムに弾力性を構築するものではありません。 プッシュオフ時における多大な筋力要求。筋膜弾性のリコイルを利用することでプッシュオフ時における甚大な筋肉の努力要求を減らし、動作はよりコントロールしやすくなり、より容易に少ない燃費で行うことができます 。 発見#2:筋膜システムは、反復するプログラムよりも変動するものにより反応をする。 エビデンスによると、筋膜システムは,角度、テンポ、そして負荷の様々な種類のベクトルによってより鍛えられるとされています (Huijing 2007)。一つの経路にそって筋肉(例えばエクササイズマシーンによるもの)を孤立化することは、それらの筋肉には有効かも知れませんが、周辺組織にはあまり有効ではありません。常に一方向からだけの負荷をかけるということは、人生において(ほぼ反復性のない)その身体部位に予期せぬことが起こった時、弱いものにしてしまいます。 必要なこと: 全身運動。 長い筋筋膜の鎖が働き、全身運動を取り入れることが筋膜システムを鍛えるより良い方法です。 近位からの初動。動作をダイナミックプレストレッチから(遠位の伸展)始めるのがベストですが、これに望む方向への近位からの初動を伴わせ、ゴム製の振り子のように身体の遠位部が流れに続くようにします 。 適合動作。 *パルクール(ジェームスボンドの映画、カジノロワイヤルの冒頭が素晴らしい例なので参考にしてください)のように適合が必要とされる複雑な動きに、反復性のエクササイズプログラムはかないません。 *(訳者注)パルクール(フランス発祥の運動方法。走る、跳ぶ、登るなどの移動動作で身体を鍛える方法) 必要ではないこと: 反復動作。クライアントが同じ線上で何度も繰り返し動作を行うマシーン(または思考)で、筋膜弾性を上手に築くことはできません。 常により高いレベルの負荷で実践すること。多種多様な負荷は筋膜の異なった側面を鍛えます。限界値に近い負荷のみに固執することで強化できる靭帯もありますが、他の構造が弱くなってしまいます。負荷を変化させることがより良い方法でしょう。 常に同じテンポでトレーニングすること。同様に、異なったテンポでトレーニングを行うことで、異なる筋膜構造の強度や弾力性が構築されます。 発見#3:筋膜システムは、筋肉よりもずっと多く神経支配されています。ですから固有受容感覚や運動感覚は主に筋膜にあり、筋肉ではないのです。 これは多くのフィットネスの専門家とって、発想の転換をするのが難しいコンセプトかもしれませんが、事実として:筋膜組織には筋肉の10倍にも及ぶ感覚受容器があるのです (Stillwell 1957)。 筋肉には紡錘があり、筋肉の長さ(時間の経過と共に長さの変化率)を測っています。これらの紡錘も筋膜受容器としてとらえられるとはいえ、とりあえず筋肉のものとして、譲っておきましょう(Van der Wal 2009)。 それぞれの紡錘には、およそ10個の受容器が筋外膜の表面、腱と筋膜の付着部、近隣の靭帯や浅層といった周囲の筋膜に存在します。 これらの受容器には(繊維の伸張度を計ることによって)負荷を計るゴルジ腱紡錘、圧を測るパチーニ小体終末、中枢神経系統に軟部組織の剪断力を伝えるルフィーニ終末、そして上記のもの全てに加え痛みも伝える、至る所にある小さな間質性神経終末が含まれます(Stecco et al. 2009; Taguchi et al. 2009)。 テンセグリティー 連続した部分としてではなく、全体としての筋膜システムを一度理解すると、身体は、それ自体が生き生きとしたテンセグリティーとして表現されます(「張力-統合」)(Fuller 1975)。 支柱は骨のようなもので、押し出し、筋膜のネットは紐や膜のようなもので、引き込みます。これら全てにより私たちが「形」と呼ぶものが作られます。私達の身体がこのように細胞的に、そしてマクロレベルで働いているというのが明らかになってきました(Ingber 2008)。 もちろん、我々人間のテンセグリティーは、神経系によって生命を吹き込まれ、筋肉を介してとても調整されやすいものですが、これらの構造の特性を探求していくことは、私たちの身体にとって価値のあることなのです。 ですから、筋肉が動いていると感じると述べることは、少し不適切なのです。あなたは筋肉よりも筋膜組織の声を“聞きとっている”のです。この基本的でありつつ目を見張るような事柄に関連した、3つの興味深い発見があります。 ほとんどの靭帯は、平行ではなく筋肉と連続して配列されています(Van der Wal 2009)。これはつまり筋肉が緊張すると、靭帯は、その位置がどこであろうと自動的に関節を安定させるために緊張します。靭帯は関節が最大伸展位、もしくは捻れがないかぎり機能しないという考えは、今や時代遅れなのです;例をあげると、靭帯はプリーチャーカールの動作の終了時のみでなく、動作の全行程で機能しています。 神経終末の配列は遺伝的なものでもなく、間違いなく私達が筋肉と呼ぶ解剖学的区分けによるものでもなく、その個人の、その位置に加わる力によって配列されます。動作脳の中には「三角筋」という表示はありません。それは単に大脳皮質にあるコンセプトであり生物学的組織に存在するものではないのです。 明らかに関節靭帯の受容器よりも、皮膚やその近辺にあるセンサーはより活発に動作を感知し、調整しています(Yahia, Pigeon & DesRosiers 1993)。 必要なこと: 皮膚と表面組織への刺激が固有感覚受容を高める。筋膜の固有感覚受容を高めるには、皮膚や表面組織をさすったり、動かしたりすることが重要です。あるウエイトリフターは、競技の前に野菜ブラシで体をこすり良い結果を納めています。 筋膜組織を感じるように、クライアントを導く。自分自身及び、そのクライアントが、筋肉ではなく周辺筋膜組織に注意を払うことで怪我を予防でき、運動感覚の認知がより正確にしっかりと伝達されます。高いレベルの鋭敏な運動感覚(猫を思い浮かべてください)を伴った感覚的な身体活動は、タフであることよりも、より良く怪我を予防することができるでしょう。 必要ではないこと: 孤立した筋肉の方向性。一つの筋肉やそのグループだけのエクササイズをするということは、ほぼ不可能です;全てのエクササイズにおいて複数の神経が刺激され、複数の筋肉が関わり、 “上流”“下流”を含め動作が行われている周辺、全ての筋膜組織が動員されているのです。 関節受容器の強調:時に靭帯は筋肉により緊張しますが、関節受容器を強調することは(重要ではあるものの)皮膚からその下の全領域にかけて、より全体に注意を向けるという考えに変える必要があります。 この議論は生体力学の要素に重点を置いてきました;栄養学、体液の考察、近年研究題材となっている筋膜の異なった構成などに関しては省略されています。トレーニングにおける筋膜の役割をより深く理解することは、私たちの知覚、ワーク、言語や効果に変化をもたらします。筋膜は、単にぴったりとくっつくサランラップのようなものではないのです。 参考文献 Chen, C.S., et al. 1997. Geometric control of cell life and death. Science, 276 (5317), 1425–28. Chino, K., et al. 2008. In vivo fascicle behaviour of synergistic muscles on concentric and eccentric plantar flexion in humans. Journal of Electromyography and Kinesiology, 18 (1), 79–88. Desmouliere, A., Chapponier, C., & Gabbiani, G. 2005. Tissue repair, contraction, and the myofibroblast. Wound Repair Regeneration, 13 (1), 7–12. Fascia Congress. 2009. www.fasciacongress.org/2009. Fuller, R.B. 1975. Synergetics. New York: Macmillan. Grinnell, F. 2008. Fibroblast mechanics in three-dimensional collagen matrices. Trends in Cell Biology, 12 (3), 191–93. Grinnell, F., & Petroll, W. 2010. Cell motility and mechanics in three-dimensional collagen matrices. Annual Review of Cell and Developmental Biology, 26, 335–61. Horwitz, A. 1997. Integrins and health. Scientific American, 276, 68–75. Huijing, P. 2007. Epimuscular myofascial force transmission between antagonistic and synergistic muscles can explain movement limitation in spastic paresis. Journal of Biomechanics, 17 (6), 708–24. Huijing, P.A., & Langevin, H. 2009. Communicating about fascia: History, pitfalls and recommendations. In P.A. Huijing et al. (Eds.), Fascia Research II: Basic Science and Implications for Conventional and Complementary Health Care. Munich, Germany: Elsevier GmbH. Iatrides, J., et al. 2003. Subcutaneous tissue mechanical behaviour is linear and viscoelastic under uniaxial tension. Connective Tissue Research, 44 (5), 208–17. Ingber, D. 1998. The architecture of life. Scientific American, 278, 48–57. Ingber, D. 2008. Tensegrity and mechanotransduction. Journal of Bodywork and Movement Therapies, 12 (3), 198–200. Kubo, K., et al. 2006. Effects of series elasticity on the human knee extension torque-angle relationship in vivo. Re- search Quarterly for Exercise & Sport, 77 (4), 408–16. Langevin, H. 2006. Connective tissue: A body-wide signaling network? Medical Hypotheses, 66 (6), 1074–77. Langevin, H., et al. 2010. Fibroblast cytoskeletal remodeling contributes to connective tissue tension. Journal of Cellular Physiology. E-pub ahead of publication. Oct. 13, 2010. Myers, T.W. 1998. Kinesthetic dystonia. Journal of Bodywork and Movement Therapies, 2 (2), 101–14. Myers, T.W. 2009. Anatomy Trains: Myofascial Meridans for Manual and Movement Therapists. New York: Churchill- Livingston. Sbriccoli, P., et al. 2005. Neuromuscular response to cyclic loading of the anterior cruciate ligament. The Amercian Journal of Sports Medicine, 33 (4), 543–51. Schleip, R. 2003. Fascial plasticity—a new neurobiological explanation. Journal of Bodywork and Movement Therapies. Part 1: 2003, 7 (1), 11–19; part 2: 2003, (2), 104–16. Shacklock, M. 2005. Clinical Neurodynamics. Burlington, MA: Butterworth-Heinemann. Shultz, L., & Feitis, R. 1996. The Endless Web. Berkeley, CA: North Atlantic Books. Snyder, G. 1975. Fasciae: Applied anatomy and physiology. Kirksville, MO: Kirksville College of Osteopathy. Stecco, C., et al. 2009. Treatment of phantom-limb pain according to the fascial manipulation technique: A pilot study. In P.A. Huijing et al. (Eds.), Fascia Research II: Basic Science and Implications for Conventional and Complemen- tary Health Care. Munich, Germany: Elsevier GmbH. Stillwell, D.L. 1957. Regional variations in the innervation of deep fasciae and aponeuroses. The Anatomical Record, 127 (4), 635–53. Taguchi, T., et al. 2009. The thoracolumbar fascia as a source of low back pain. In P.A. Huijing et al. (Eds.), Fascia Re- search II: Basic Science and Implications for Conventional and Complementary Health Care. Munich, Germany: El- sevier GmbH. Van der Wal, J. 2009 The architecture of the connective tissue in the musculoskeletal system: An often overlooked functional parameter as to proprioception in the locomotor apparatus. In P.A. Huijing et al. (Eds), Fascia Research II: Basic Science and Implications for Conventional and Complementary Health Care. Munich, Germany: Elsevier GmbH. Varela, F., & Frenk, S. 1987. The organ of form. Journal of Social and Biological Structures, 10 (1), 1073–83. Yahia, L.H., Pigeon, P., & DesRosiers, E.A. 1993. Viscoelastic properties of the human lumbodorsal fascia. Journal of Biomedical Engineering 15, 425–29.

筋膜の弾性を向上させる(ビデオ)
10月3日に、東京で開催された、トーマス・マイヤースのセミナー”筋膜のフィットネス”のクラス風景から、2つのムーブメントのアイデアを収録。トムが、筋膜の持つ弾力性を活かして、更にゴムのように弾む身体を作るために、簡単にできる動きのアイデアや、型にとらわれることなく、自分自身の身体が動きたいように、自由に探検してみることの提案をシェアしてくれています。

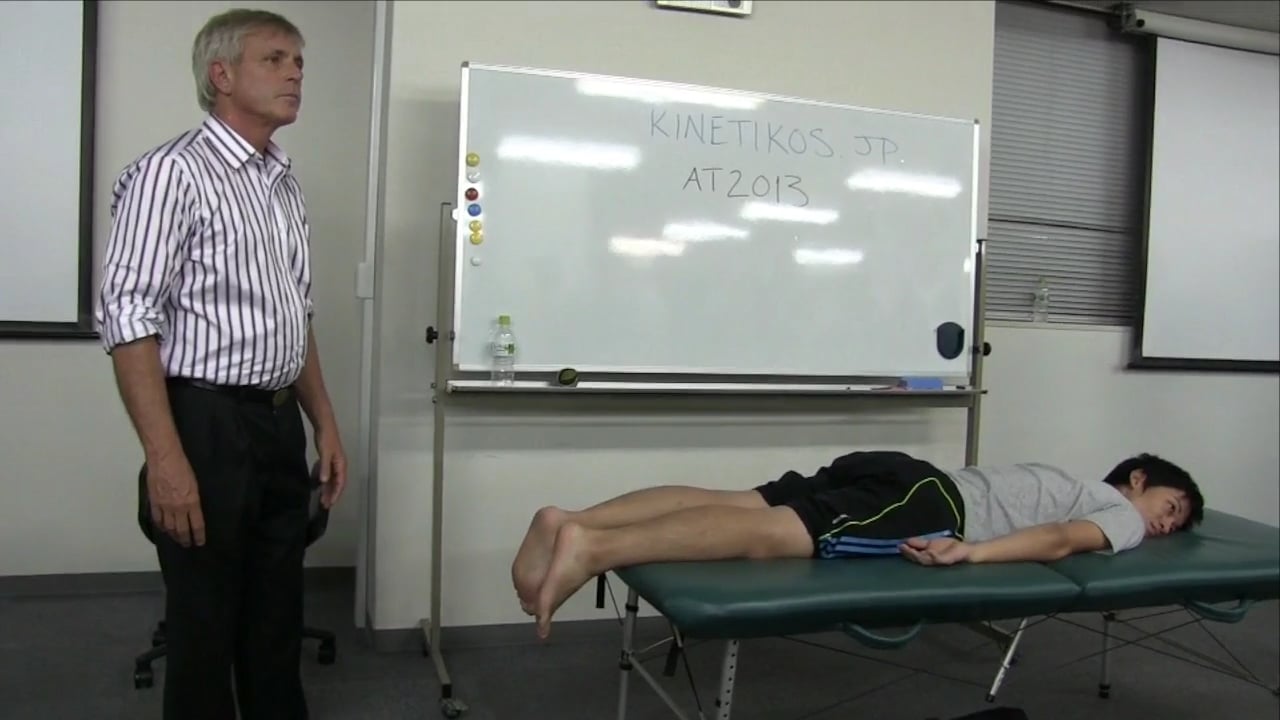
構造の支点 ポイント1/7(ビデオ)
(ポイント2/7はこちらへ) 先週、10月4日&5日に東京で開催したトーマス・マイヤースのハンズオンセミナー”アナトミートレイン:構造の支点”の7つの重要なポイントの中から、まずは1つめのポイントのご紹介をします。構造の一番下部に位置する、このポイントは、私達のグラウンディングに深く影響します。 ポイント #1:地面 - 後脛骨筋遠位付着部 ポイントを見つける:距舟関節の下方、内側アーチの中間 方向:足を内顆の方向に向かって上方へ 重要性/意義:コアの底部 - ディープフロントライン全体を目覚めさせ、アーチ、背骨の 前部、骨盤底にも影響あり
構造の支点 ポイント2/7(ビデオ)
(ポイント1/7はこちらへ) (ポイント3/7はこちらへ) 10月4日&5日の2日間にわたって開催された、トーマス・マイヤースのハンズオンクラス“構造の支点”の7つの重要ポイントの中から、2つ目のポイントである、腓腹筋、ヒラメ筋、アキレス腱の接合点へのハンズオンテクニックをご紹介します。 ポイント #2:不安感 -腓腹筋/ヒラメ筋/アキレス腱の接点 ポイントを見つける:腓腹筋の内側頭と外側頭の間の区分の下端 方向:下腿部に向かってまっすぐ前方に 重要性/意義:前方突出の一番の被害者 - スーパーフィシャルバックライン全体に影響する。トリ ートメントは骨盤を中心とし、しばしば不安感を減少させる効果をもつ。

構造の支点 ポイント3/7(ビデオ)
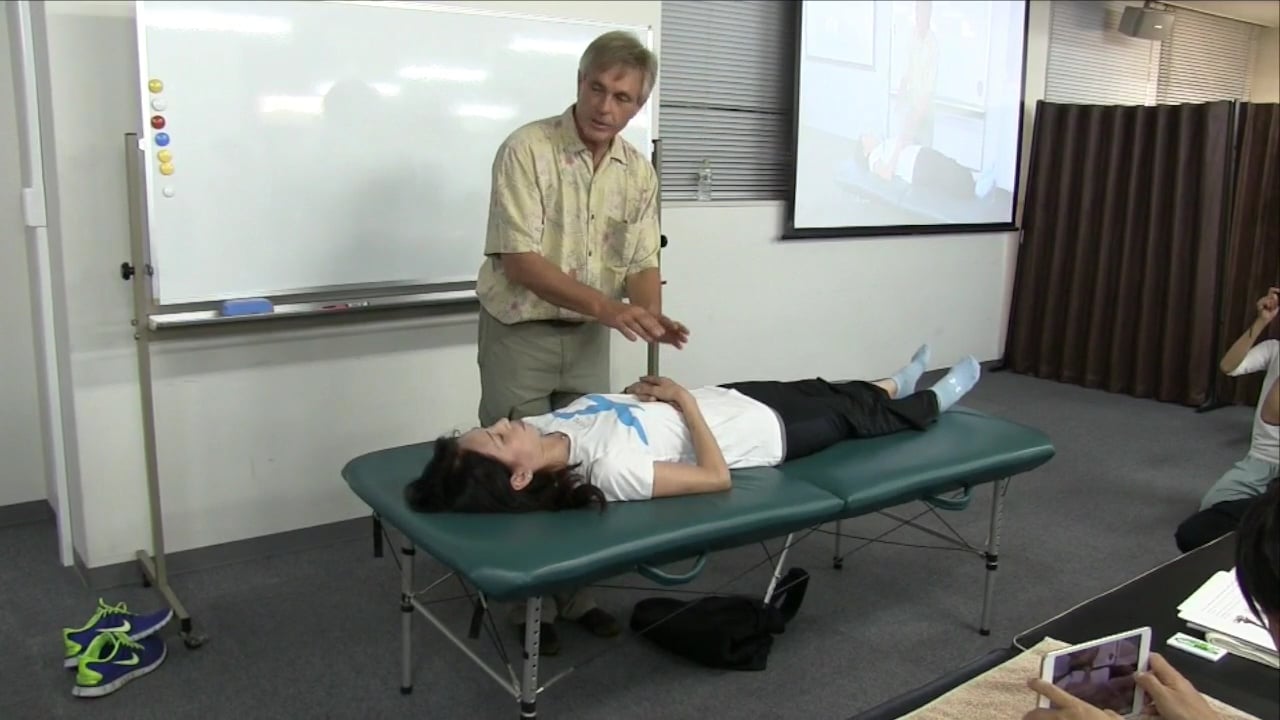
(ポイント2/7はこちらへ) (ポイント4/7はこちらへ) 10月4日&5日に開催された、トーマス・マイヤースのハンズオンクラス”構造の支点"の7つの重要ポイントの3番目、小転子へのアプローチの手技解説の模様をお送りします。小転子に直接アプローチするためには、組織が既にオープンになっている必要がありますが、股関節内側の組織をリリースして脚を股関節から引き出してくるようなテクニックもご紹介しています。 ポイント #3:後退 - 小転子 股関節の内側 ポイントを見つける:薄筋と大内転筋の間、座骨枝後部のすぐ下側 方向:股関節の内側に向かって上方、外側へ 重要性/意義:テキストに書かれている最高の“脚長伸ばし”大腿骨の股関節への後退を減少させる
構造の支点 ポイント4/7(ビデオ)
(ポイント3/7はこちらへ) (ポイント5/7はこちらへ) 10月4日&5日に開催された、トーマス・マイヤースのハンズオンクラス ”構造の支点” の7つの重要ポイントの4番目、仙腸関節へのアプローチの手技の模様をお届けします。仙腸関節のエリアには多くの靭帯が存在します。結合組織が密な部分であればある程、固有受容器の密度も高くなる、つまりこのエリアへのアプローチはパワフルなものになり得るのです。 ポイント #4:前に踏み出す - 仙腸関節 ポイントを見つける:上後腸骨棘(PSIS)の内側 方向:仙腸靭帯背側に向かって前方、外側へ 重要性/意義:脚に対する背骨の“モーターマウント“バランスのとれた成熟した歩行の鍵

構造の支点 ポイント5/7(ビデオ)
(ポイント4/7はこちらへ) (ポイント6/7はこちらへ) 10月4日&5日に開催された、トーマス・マイヤースのハンズオンクラス”構造の支点”の7つの重要ポイントの5つ目。ミッドドーサルヒンジ=中背部のヒンジのポイントへのアプローチをご紹介します。T6~7の両脇から肋椎関節方向に向かってのアプローチで、身体前面のラインへ影響を与えます。 ポイント #5:保護 ‒ 中背部ヒンジ ポイントを見つける:T6-7の両脇、中心線から約3cm離れたところ 方向:肋椎関節及び肋横突関節に向かって前方、やや外側 重要性/意義:身体の前側(スーパーフィシャルフロントライン、ディープフロントライン)をオ ープンにし、呼吸を楽にし、胸椎過度後弯を減少させる

構造の支点 ポイント6/7(ビデオ)
(ポイント5/7はこちらへ) (ポイント7/7はこちらへ) 2013年10月4日&5日に開催された、トーマス・マイヤースのハンズオンクラス”構造の支点”の7つの重要ポイントの6つ目。胸骨と鎖骨が出会う、鎖骨内側の下側、第一肋骨に近い胸鎖接合へのアプローチをご紹介します。肩の動きをオープンにしてリストアする重要なポイントです。 ポイント #6:リーチアウト -胸鎖接合 ポイントを見つける:鎖骨内側の下側、第一肋骨に近いところ 方向:後方、内側からスタートして外側へ 重要性/意義:スーパーフィシャルフロントアームライン及びディープフロントアームラインを通 して、肩の動きをオープンにし、リストアする